ПРОТИВОВОСПАЛИТЕЛЬНОЕ ДЕЙСТВИЕ ИФН-a В НЕБОЛЬШИХ ДОЗАХ
– ЛАБОРАТОРНОЕ ИССЛЕДОВАНИЕ ЛЕЙКОЦИТОВ У СВИНЕЙ
Др. Массимо Амадори доктор медицинских наук, ветеринар, Брешиа, Италия
КРАТКОЕ ИЗЛОЖЕНИЕ
Взаимодействие мононуклеаров периферической крови (МНПК) и ИФН–α малой концентрации вызывает сильное,зависящее от дозы снижение экспрессии CD14.
Стимулированное бактериальными липополисахаридами (ЛПС) снижение аккумуляции ФНО- α в МНПК свиней и альвеолярных макрофагах (АМ) является причиной уменьшения концентрации ИФН-α в 10 раз (0.5 единиц мл-1). При этом экспрессия ФНО- α гена в АМ также значительно сокращается в отличие от клеток реакцией в 50 единиц мл-1 ИФН-α. Напротив, экспрессия IL-1β и IL-6 стимулируется в обоих случаях. Таким образом, ИФН-α играет важную роль в контроле противовоспалительного ответ на бактериальные эндотоксины у свиней.
Ключевые слова: небольших концентраций ИФН-α, воспаление, лабораторное исследование.
ВВЕДЕНИЕ
В ходе интенсивной репродуктивной деятельности свиньи страдают от различных заболеваний (в послеотъемный период), среди которых СРРС (свиной репродуктивный и респираторный синдром) и СМПИ (синдром мультисистемного послеотъемного истощения поросят), вызванных соответственно PRRS-вирусом и цирковирусом свиней. Несмотря на распространенность вирусных инфекций при размножении свиней, только больные животные подвергаются воспалительному действию и проявлению нарушений функционирования иммунной системы.
Фактически, вышеупомянутые вирусные инфекции способствуют распространению воспалительных процессов при высокой концентрации бактериальных липополисахаридов в период размножения.
Принимая во внимание данную гипотезу, мы сфокусировали основное внимание на пероральном лечении с помощью ИФН-α, проявившем эффективность в терапии различных экспериментальных моделей хронического воспаления и аутоиммунных заболеваний.
В сущности, согласно с опубликованными результатами потенциальная роль ИФН-α в регулировании врожденного и приобретенного иммунного ответа у многих видов животных, включая свиней, очевидна. Следовательно, лабораторное исследование было посвящено воздействию ИФН-α на лейкоциты свиней в процессе реакции на бактериальные ЛПС.
МАТЕРИАЛЫ И МЕТОДЫ
КЛЕТКИ
Образцы гепаринизированной венозной крови, взятые у отъемных поросят (средний вес 30 кг), в разведении 1:2 в средеRPMI 1640 были дополнены антибиотическим воздействием и очищены с помощью Histopaque® – 1077. Полученные МНПК были дважды обработаны RPMI 1640, рассчитаны и культивированы на 2 х 106 мл-1 в RPMI 1640 + 10% фетальной бычьей сыворотки (ФБС) на поверхности для культивирования клеток с 12 лунками.
АМ были взяты у отъемных поросят, умерщвленных после общей анестезии (бронхоальвеолярный лаваж). Культивирование клеток проходило по предыдущему принципу (RPMI 1640 + 10% ФБС) на поверхности для культивирования клеток с 12 лунками.
Образцы альвеолярных макрофагов и крови были собраны под контролем ветеринара и органов здравоохранения.
ИНТЕРФЕРОН-α
Очищенный человеческий ИФН-α, полученный из лимфобластоидных клеток, был заморожен (-80˚С) в определенных количествах и использовался в рассматриваемых лабораторных экспериментах. Биологическая активность определялась торможением цитопатического эффекта интерференцией вируса везикулярного стоматита бычьих MDBK клеток и человеческих HEP-2 клеток.
ЛЕЧЕНИЕ С ПРИМЕНЕНИЕМ ИФН- α И ЛПС
К свиным МНПК и АМ, культивированным на поверхности с 12 лунками, был добавлен ИФН-α разной концентрации (0-50 единиц мл-1) для дальнейшего культивирования при температуре 37˚С во влажной камере (СО2 – 5%). По прошествии 16-24 часов клетки стимулировали с помощью 1 μg мл-1 бактериальных ЛПС кишечной палочки 0111:B4, либо оставляли как контрольный образец. В последствии клетки были повторно культивированы (37˚С, 5% СО2). В ходе этого клетки проходят проточную цитометрию МАТ для CD14 и воздействие ревертазы для генов основных воспалительных цитокинов; культивационная среда проходит биоанализ на наличие фактора некроза опухоли (ФНО-α).
По 10 единиц лимфобластоидного ИФН человека на килограмм веса (добавленного в пищу) применяли для 4 свиней (через 30 дней после рождения) на протяжении 15 дней в лиофилизированной форме. 5 остальных свиней того же возраста и генетического профиля использовались как контрольный образец. На 15-ый день перорального применения ИФН-α были взяты образцы крови и помещены в гепаринизированные пробирки. Уровень концентрации используемого ИФН-α был намного ниже обычного уровня в сыворотке или кишечной секреции зараженных вирусом свиней.
ПРОТОЧНАЯ ЦИТОМЕТРИЯ
МНПК и АМ были получены с использованием ЭДТК (0.02%). Затем к ним добавили неприлипающие клетки, дважды обработали фосфатно-солевым буферным раствором (ФСБ, рН 7.4) и оставили в буфере для проточной цитометрии (ФСБ, рН 7.4, 2% ФБС – для термальной обработки, азид натрия 0.1%).
Анализ клеток проводился с помощью цитометра Bryte-
HS (BIO-RAD) для поверхностной экспрессии CD14 с моноклональными антителами (MIL-2). На селекционной сетке были проанализированы 10 000 клеток. Процент CD14+клетки был получен путем субтракции количества, соответствующего обозначенным только мышиным анти-иммуноглобулином на флуоресцеин.
ПЕРЕМЕЩЕНИЕ ОБРАБОТАННЫХ ИФН-α КЛЕТОК
Свиные МНПК культивировались по 2 х 10 мл-1 на 2 поверхностях для культивирования с 12 лунками (донорные и рецепиентные соответственно). В 6 донорских лунок (по две) добавили по 0, 0,5 и 50 единиц мл-1 ИФН-α.
– По прошествии 4 часов инкубации (при 37˚С и 5% СО2) из 3 лунок (по одной обработанной ИФН-α) изъяли неприлипающие клетки, которые дважды обработали RPMI 1640 с ФВС (10%).
2 х 105 обработанных ИФН-α клеток были впоследствии перенесены в 3 реципиентные лунки для дальнейшего культивирования на период ночи (37°C, 5% CO2) вместе с контрольными клетками в остальных реципиентных лунках. На следующий день все клетки подверглись стимулированию 1 μg мл-1 бактериальных ЛПС на 16 часов.
БИОАНАЛИЗ ФНО-α
Образцы среды пребывания обработанных ИФН-α и/или ЛПС брались на всех фазах эксперимента и замораживались (при температуре -80°C) для проведения биоанализа на ФНО-α. Ту же процедуру проделывали и для МНПК, в которые добавляли изогенные клетки, обработанные ИФН-α.
Клетки WEHI 164 (ATCC) в среде RPMI 1640 с 10% ФБС (3 х 104 на лунку) были помещены на поверхность с 96 лунками при наличии последовательно разведенных образцов по log2 и 1 μg мл-1 актиномицина D. По прошествии инкубационного периода продолжительностью 18 часов (при 37°C) с помощью 3-(4,5-диметилтиазол-2-xl)2,5-дифенил-тетразол-бромид было определено наличие жизненно важных клеток.
– Концентрацию ФНО-α определяли способом сопоставления конечных образцов со стандартным препаратом рекомбинантного свиного ФНО-α.
ПОЛУКОЛИЧЕСТВЕННЫЙ RT–PCR АНАЛИЗ АМ
Согласно инструкции РНК была изолирована от АМ триазолом. Через стандартные процедуры 3 μg РНК инверсивно транскрибировалась выборочными гексамерными РНК-затравками и ревертазой вируса мышиного лейкоза Молони (ВМЛМ).
Амплификация 6-оксипурин-фосфорибозилтрансферазы было необходимо для нормализации содержания кДНК в различных образцах клеток.
Для проведения амплификации были созданы соответствующие условия: денатурация при 94°C на 5 мин, сопровождающаяся 25 аналогичными циклами продолжительностью 30 секунд при 94°C, 30 секунд при 56°C, 45 секунд – при 72°C + 3 минуты при той же температуре. Для завершения ПЦ-реакции в каждом ген-мишени определенное количество кДНК разводили с ПЦР-буферном растворе (50 мМ KCI, 10 мМ Трис-HCl, 2 мМ MgCl2) до 50 μl – 50 пикомольРНК-затравки, вместе с дНТФ (0,8 мМ полученной концентрации) и 2 единицы ДНК-полимераза Taq. Экспрессия генов цитокинов исследовалась через ко-амплификацию ген-мишеней и ГПРТ при аналогичных условий цикла ПЦР.
Уровень экспрессии генов был определен посредством определения соотношения ген-мишени и ГПРТ, используя программу IMAGEQUANT. Тесты проводились по трем схемам для дальнейшего статистического анализа.
Использовались следующие РНК-затравки (праймеры):
- – ГПРТ передний праймер: 5’ – CCATCACATCGTAGCCCTCT – 3’
- – ГПРТ обратный праймер: 5’ – TCAAATCCAACAAAGTCTGGC –3’
- – IL-1β передний праймер: 5’ – ATGGCCCCAAAGAGATGAAG – 3’
- – IL-1β обратный праймер: 5’ – TGCACGTTTCAAGGATGATG – 3’
- – ФНО-α передний праймер: 5’ – AACCTCAGATAAGCCCGTCG – 3’
- – ФНО-α обратный праймер: 5’ – ATGGCAGAGAGGAGGTTGAC – 3’
- – IL-6 передний праймер: 5’ – TGCTCTTCACCTCTCCGGAC – 3’
- – IL-6 обратный праймер: 5’ – CTTTCTGGAGGTAGTCCAGG – 3’.
СЕКРЕЦИЯ СУПЕРОКСИД-АНИОНА
По прошествии инкубационного периода (ночь) с 0 и 0.5 единиц мл-1 ИФН-α секреция супероксид-аниона в среде культивирования АМ измерялась с помощью анализа стимуляции цитохрома С форбол-12-миристат-13-ацетатой (ФМА).
РЕЗУЛЬТАТЫ
► Супрессирующий эффект экспрессии CD14
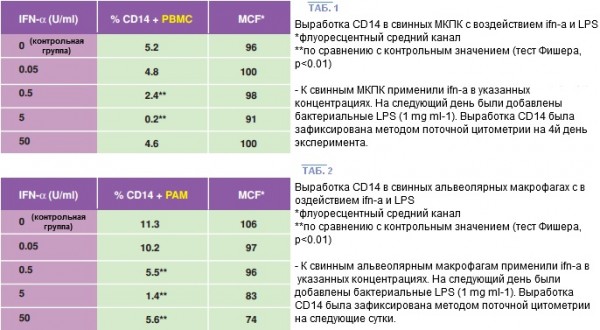
В ходе 4 отдельных экспериментов было определено, что воздействие ИФН-α уменьшает наружную экспрессию CD14 в МНПК (5 животных) и в АМ (1 животное) зависимости от дозы. Пиковая активность была зафиксирована при 5 единицах мл-1. Интересно заметить, что такая активность не изменилась и после добавления ЛПС.
► Уменьшение секреции ФНО-α
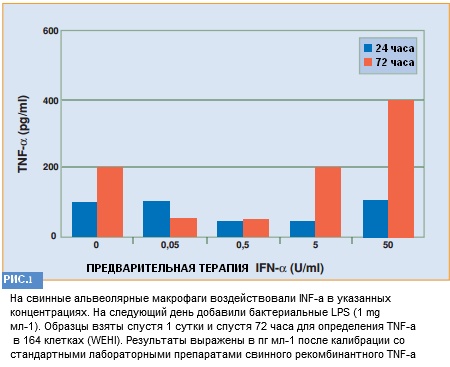
После воздействия на бактериальный ЛПС секреция ФНО-α в АМ в последствии обработки ИФН-α неизменно снижается (в 2-4 раза) во всех 4 экспериментах.
По результатам сравнения экспериментов на экспрессию CD14 наибольшее снижение ФНО-α в среде культивирования наблюдалось при в 10 раз меньшей концентрации либо при 0.5 единиц мл-1 ИФН-α.
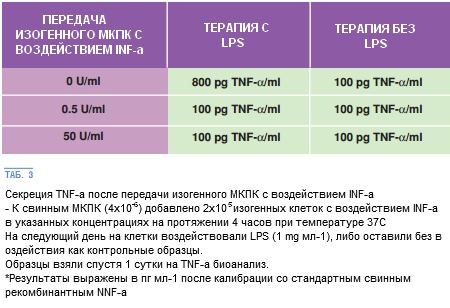
В МНПК уровень секреции ФНО-α после добавления ИФН-α в двух экспериментах значительно не менялся. Несмотря на это, МНПК существенно повлияли на снижение секреции ФНО-α впоследствии добавления изогенных клеток, обработанных ИФН-α, в дальнейших экспериментах. Более того, МНПК у 3 из 4 подопытных свиней вырабатывали значительно меньшее количество ФНО-α после проведения в естественных условиях ЛПС-стимуляции (<50 pgs мл-1 по сравнению с >200 pgs мл-1 у всех 5 свиней) (t 2.98; p < 0.05).
► Экспрессия генов цитокинов
В трех отдельных ОТ-ПЦР экспериментах на АМ ИФН-α 0.5 единиц мл-1 (в отличие от ИФН-α 50 единиц мл-1) существенно уменьшил экспрессию гена ФНО-α после стимуляции с помощью бактериального ЛПС. С другой стороны при тех же условиях экспрессия гена IL-1β до разного уровня стимулировалась ИФН-α концентрацией как 0.5, так и 50 единиц мл-1. Существенных изменений экспрессии гена IL-6 выявлено не было.
► Выделение супероксид-аниона
В ходе 2 разных экспериментов профилактика АМ с помощью ИФН-α (0.5 единиц мл-1) способствовала 3-кратной редукции секреции супероксид-аниона (OD550) после стимуляции форбол-12-миристат-13-ацетатой (ФМА).
 КОММЕНТАРИЙ
КОММЕНТАРИЙ
До недавнего времени оставалось неизвестным антивирусное, иммуностимулирующее и антипролиферативное действие ИФН-α, особенности определения необходимых концентрации и времени применения. Интересно заметить, что по сравнению с антивирусным воздействием кривая силы ответа от дозы ИФН-α зачастую колоколообразная – по функции модулирования иммунной системы: выше дозволенного уровня концентрации биоактивность падает, а иногда даже уходит в минус (напр. сравнение стимуляции и супрессии первичного ответа антитела на овечьи эритроциты у мышей).
В частности, эффективность стимуляции иммунной системы, используя низкие либо средние концентрации, явно преобладает над показателями кривых традиционного ответа на дозу ИФН-α, благодаря его гормонной природе и типу действия вестественных условиях (аутокринное/паракринное).
Данное предположение полностью подтверждают полученные после перорального применения ИФН-α малых доз результаты лечения различных инфекций и аутоиммунных заболеваний.
Вырабатываемый ИФН-α сигнал передается через систему амплификации, индуцируется малейшими пероральными дозами (1-10 IU/кг массы тела) и исчезает после применения бóльших ежедневных доз.
Бóльшие дозы вызывают провоспалительный ответ Тh-1, в то время как парэнтеральные инъекции больших доз могут вызвать воспаление и пиретическую реакцию у людей и у животных.
В экспериментах со свиньями модели PRRS и PMWS оказались полезными для определения вероятной роли ИФН-α в контролировании воспалительных реакций.
Фактически, переход от вирусной инфекции до болезни связан со снижением секреции эндогенного ИФН-α и основного вырабатывания IL-10. Таким образом, роль ИФН-α как регулятора воспалительных реакций может оказаться весьма существенной, т. к. вирус PRRS значительно повышает предрасположенность свиней к бактериальному ЛПС. Более того, последовательность ДНК второго типа свиного цирковируса (связанного с PMWS) способствует искусственной транскрипции генов ИФН-α. Важным является то, что пероральное применение генов ИФН-α отъемными свиньями вызывает снижение уровня потерь (от болезненности, истощения, смерти) при поражении PRRS и/или PMWS в период размножения. Допускаемая роль ИФН-α в качестве модулятора воспалительного ответа полностью соответствует результатам данного исследования; в частности, супрессирующее действие CD14 (клеточный рецептор ЛПС/ЛПС-связывающего комплекса) показывает, что чувствительность к бактериальным эндотоксинам может существенно модифицироваться с помощью ИФН-α.
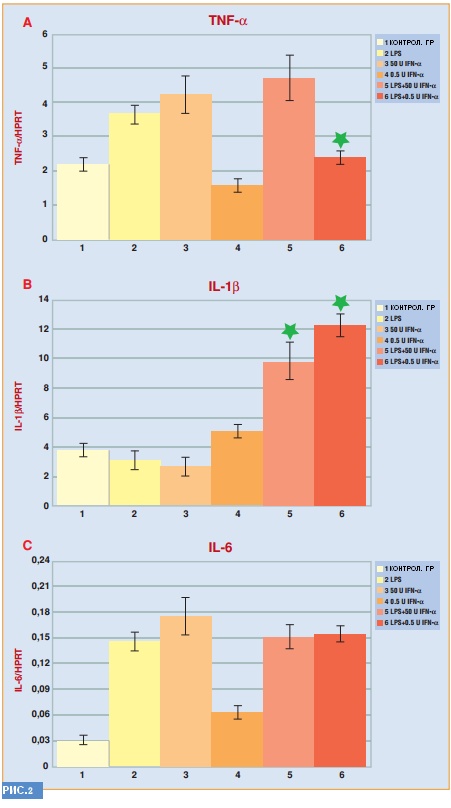

Пониженная секреция ФНО-α, отмеченная в свиных АМ, относится к присутствию небольшой концентрации ИФН-α до применения ЛПС. Подтверждение регуляторного действия обработанных ИФН-α клеток является чрезвычайно важным: такие клетки «провоцируют» реципиентные изогенные клетки на супрессирующий эффект ответа ФНО-α на бактериальные ЛПС.
Не менее существенно снижение ФНО-α-ответа на PBMC у свиней вследствие перорального применения ИФН-α: внимание акцентируется на амплификации регулирующего сигнала в естественных условиях. Также было отмечено абсолютное сходство между концентрацией ИФН-α и ФНО-α-реакцией – в протеин и ген-экспрессии АМ после применения ЛПС.
Эффективность адаптивного иммунного ответа не уменьшается при действии малых доз ИФН-α, что явно прослеживается при экспрессии генов IL1-β и IL1-6. К тому же, повышенная экспрессия гена IL1-β выдается безвредной, благодаря посттрансляционным контрольным системам (в большинстве случаев эпителиальный рецептор и IL1 рецептор антагонист).
Так как подобных контрольных систем для ФНО-α нет, его ген и цитокин рассматриваются как важнейшие и неотъемлемые показатели низкого титра ИФН-α для контроля острого воспалительного ответа.
Больше того, интерес для исследователей представляет процесс быстрого супрессирующего эффекта CD14 при низкой концентрации ИФН-α: индуцированный ЛПС/ЛПС-связывающим комплексом внутриклеточный сигнал амплификации может быть снижен, высвобождение свободного CD14 – вызвать значительное фагоцитарное действие на бактериальный ЛПС.
– В контексте защитной функции ИФН-α можно рассматривать как фактор гомеостатического контроля. С этого можно сделать вывод, что чувствительность лейкоцитов у свиней к бактериальным эндотоксинам снижается при применении малых доз ИФН-α. Этот факт несомненно является действенным и для естественных условий. В случае поражения вирусными и бактериальными инфекциями ИФН-α высокой концентрации может изначально способствовать воспалительному ответу; с другой стороны, ИФН-α малых доз уменьшит его в последующих фазах снижения врожденного и приобретенного иммунного ответа – как и в случае исследованного физиологического ИФН-α-ответа.
Таким образом, кроме общеизвестных функций иддукции/регуляции врожденной иммунной системы и антивирусного действия, ИФН-α используется в более широком диапазоне, механизме гомеостатической регуляции.

